Product Overview
Midbrain dopamine (DA) neurons are critical for directing fundamental brain functions such voluntary movement, reward processing, and working memory. The substantia nigra (SN) and the ventral tegmental area (VTA) have the highest populations of DA neurons in the midbrain. The degeneration of DA neurons within the pars compacta region of the SN is a pathological hallmark of Parkinson’s disease (PD) and Lewy body dementia (LBD)[1] . For many years, powerful experimental model organisms like the mouse, fruit fly, and baker’s yeast have been used to study neurodegenerative diseases, providing insights into disease mechanisms like pathological aggregation of key proteins, the nature and processes of neuronal damage, the role of genetic determinants, and the contribution of neuroinflammation in fueling neuronal loss [2,3]. However, it appears that the use of these models has only partially elucidated some elements of the illnesses, impeding a meaningful translation into new treatments, diagnostics, and prevention.

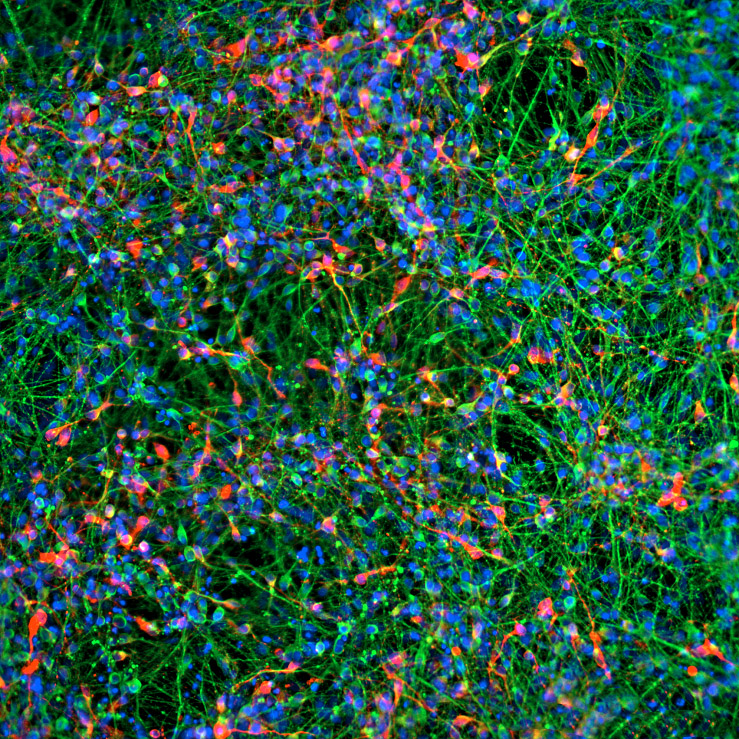
Figure 1. Human iPSCs derived dopaminergic neurons show expression of characteristic biological markers. (A) Immunostaining shows the expression of midbrain dopaminergic neuron markers FoxA2 and TH, 28 days post-thawing. (B) Quantifications for % of dopaminergic neurons, positive for the TH marker over multiple time-points. Results are expressed as means ± SEM. (C) Flow cytometry measurements demonstrate a highly specific population of fully differentiated midbrain dopaminergic neuron (TH). (D)(E) Immunostaining shows the expression of neuron marker Tuj1 (D) 21 days post-thawing and MAP2 (E) 7 days post-thawing and the absence of the proliferative progenitor marker Ki67. (F) Flow cytometry measurements demonstrate a highly pure population of fully differentiated neurons (MAP2). Nuclei were counterstained with DAPI. Scale bars, 200 µm.
iXCells Biotechnologies is proud to provide fully differentiated and functional human iPSC-derived DA neurons that display typical neuronal morphology and express all key markers of DA neurons, e.g., TH, FoxA2 (Figure 1) when cultured in the Human Dopaminergic Neuron Maturation Medium (Cat# MD-0105-100ML). In addition, our iPSC-derived DA neurons can also be co-cultured with glial cells or other cell types for drug screening platforms. Moreover, functional assays revealed that these neurons display calcium transient activity (Figure 2) and whole cell patch clamp revealed that over 75% of the neurons exhibited mature spiking, and over 50% of the neurons had spontaneous activity at a holding potential of -45 mV (Figure 3) indicating the presence of a highly mature population of neurons.

Figure 2. (A) Live single cell calcium activity was recorded using Cal-Bryte-590 fluorescent calcium indicator and imaged on the IC200 KIC® (Kinetic Image Cytometer) (Vala Sciences) and analyzed using CyteSeer® (Vala Sciences). (B) Individual Calcium Traces are color coded with respective arrows. (C) Event Frequency, maximum and mean peak amplitude, mean peak area are compared between iXCells dopaminergic neurons (red, Cat# 40HU002), iXCells cortical neurons (blue, Cat# 40HU-009) and competitor X glutamatergic neurons (grey). The results are graphed as mean ± SEM.

Figure 3. (A) An image of the cells during the experiment. The patch pipette is visible on the left. (B) The Sodium/potassium currents were recorded in voltage clamp mode with test potentials of -100 mV to 90 mV. (C) Excitatory postsynaptic currents (EPSCs) were recorded in voltage clamp mode while clamping the cell at -60 mV. (D) Evoked action potentials were recorded in current clamp mode starting with a current injection 12 pA below what is needed to hold the neuron at – 60 mV and with 3pA current steps. (E) Spontaneous activity was recorded in current clamp mode with a current injection needed for a membrane potential of -45 mV.
Product Details
| Tissue | Human iPSC-derived dopaminergic neurons (Normal) |
| Package Size | 1.0 million cells/vial; 2.0 million cells/vial; |
| Shipped | Cryopreserved |
| Storage | Liquid Nitrogen |
| Media | Human Dopaminergic Neuron Maturation Medium (Cat# MD-0105-100ML) Recovery Supplement (Cat# MD-0110-20μL) |
References
[1] Bloem, B. R., Okun, M. S. & Klein, C. (2021) Parkinson’s disease. Lancet 397, 2284–2303.
[2] Schulz-Schaeffer, W.J. (2015). Is Cell Death Primary or Secondary in the Pathophysiology of Idiopathic PD? Biomolecules 5, 1467-1479.
[3] Poewe, W., Seppi, K., Tanner, C. et al. (2017). Parkinson disease. Nat Rev Dis Primers 3, 17013